
[bibshow file=library.bib key_format=cite]
Background
Close your eyes, cover your ears, and do not touch anything. Even without any active afferent stimuli coming to the sensory areas, the brain is known to display a spontaneous activity whose nature and origin is still a matter of debate. Whatever the time is, your brain is constantly active and millions of neurons are emitting action potentials every millisecond. If not, this is a clear clinical sign that you are dead. This spontaneous activity, also called ongoing activity, is by definition the running activity of the brain when not ``facing" or processing particular stimuli. This is somehow the resting state of the brain, when no particular or at least no known actions are performed. The nature of this activity is correlated with behavioural states, being different during awake states and sleep. Its nature can be captured at different scales, depending on the device used to record it. For example with electro-or magneto encephalography (EEG or MEG), the frequency content of large scale brain electrical activity can be used to asses and characterize different cortical states of activity. It is usually divided into frequency bands:  rhythm, with frequencies around 8-12 Hz, can be seen as an awake resting state, when eyes are closed.
rhythm, with frequencies around 8-12 Hz, can be seen as an awake resting state, when eyes are closed. 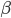 rhythm (10-30 Hz) is a kind of normal working state.
rhythm (10-30 Hz) is a kind of normal working state. 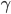 oscillations, at more than 30 Hz appear during cross-modal tasks, active and intense concentration and memory tasks (in addition to slow oscillations observed in the EEG signal, such as
oscillations, at more than 30 Hz appear during cross-modal tasks, active and intense concentration and memory tasks (in addition to slow oscillations observed in the EEG signal, such as 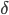 (
( 4Hz) and
4Hz) and 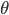 (4-10 Hz)).
(4-10 Hz)).
[youtube width="600" height="365" video_id="uhCF-zlk0jY"]
[wpcol_1half id="" class="" style=""]
Origin of Spontaneous activity
The origin of this spontaneous activity in the cortex is far from being clearly understood. Isolated neurons by themselves seem to have some homeostatic processes making them able to be spontaneously active, even if no incoming activity is present. This phenomenon has also been reported in cortical neurons [bibcite key=Llinas1991,Mazzoni2007], either taken in vitro brain slices [bibcite key=Timofeev2000] or in cultures [bibcite key=Gross1982]. Such activity, once created, can easily reverberate and be amplified in the microcircuits made by the neuronal connections, and lead to a self-sustained activity regime. How long this ongoing activity will last depends on the preparation, on the size of the network, and some other unknown parameters. Plasticity, the fact that the efficacy transmission of the synapses is not constant over time, the fact that neurons may be dying during the time of the in vitro preparation: all affect this activity, and progressively silence the network. Nevertheless, even an isolated brain displays such a spontaneous activity, i.e. the brain does not require external inputs to generate its own recurrent activity.
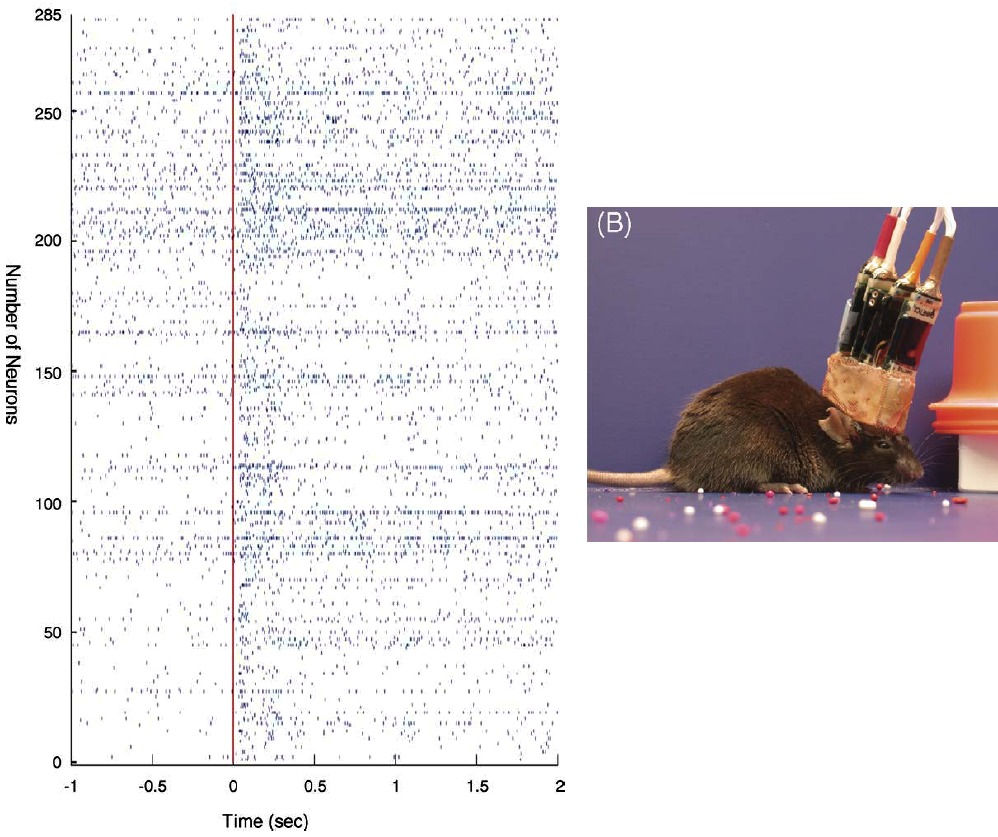
To focus on the sensory systems in vivo, the origins of ongoing activity are already found in the transduction layers, where the external world is transformed into electrical activities that are relayed to the cortex. The resting state of the brain is then an active one, and relevant information need to be extracted from these continuous and ongoing flows of sensory inputs. To illustrate this, one can look at recent data from techniques that allow to record more and more accurately the detailed and exact ongoing activity in awake animals. In Figure nearby adapted from [bibcite key=Lin2006], one can see the spiking activity in an awake mouse, and the subtle difference when the animal receives a clear external stimulation (red line). More than 250 neurons are recorded in parallel with a multi-electrode array. Understanding how pertinent information is extracted from this ongoing activity is a crucial step.
[/wpcol_1half]
[wpcol_1half_end id="" class="" style=""]
[/wpcol_1half_end]
[wpcol_1half id="" class="" style=""]
[/wpcol_1half]
[wpcol_1half_end id="" class="" style=""]
Nature of the spontaneous activity
The exact nature of the spontaneous activity is hard to capture, for several reasons. The first one is that the dynamical nature of this resting state is not that clear, and the question of its stationarity remains open. Imaging tools available nowadays (from intracellular recordings to local field potentials (LFP), voltage-sensitive dyes (VSD), two-photons) can give an insight about its nature, but not a full and exhaustive view of the electrical activity over large portions of the brain.
Age, wakefulness and anaesthetic dependence: statistics of this spontaneous activity recorded in vivo depend on the age and the state of the animal. Anaesthetics are known to perturb the balance between excitation and inhibition [bibcite key=Winters1976], leading to pathological activity that may be sometimes far from the awake regime. More and more efforts are devoted to developing awake recordings that will be as precise and controlled as in the anaesthetized context [bibcite key=Lin2006,Ferezou2006,Greenberg2008], to draw a better picture of the brain's activity. An observation that can be made from these awake recordings is that ongoing activity is rather sparse and neurons fire spontaneously at relatively low firing rates. Again the situation varies as a function of the age, the area, the species, and is used the anaesthetic, but in freely moving awake rats, sparse activity was observed intra-cellularly [bibcite key=Lee2006].
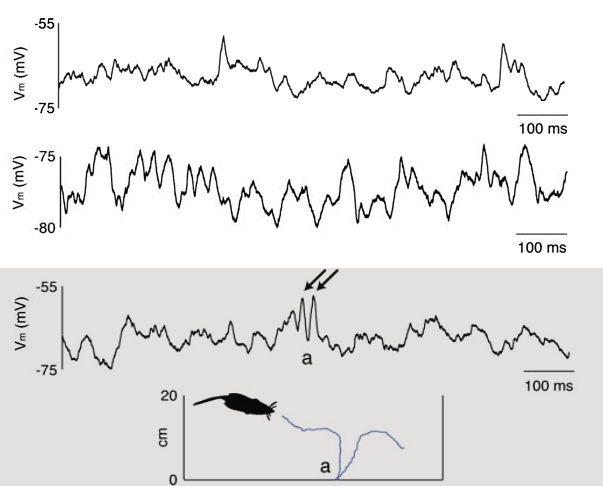
Slow Oscillations: In spontaneous activity under anaesthesia, slow oscillations observed (in the EEG but also in the membrane potential of individual neurons [bibcite key=Steriade1993a,Steriade1993]) have been considered as reflecting a switch between "up" and "down" states. In the membrane potential trace recorded in vivo, one observes some silent periods, where the membrane stays close to its resting potential (``down” state), and some very active periods where the membrane is strongly depolarized and the neuron sustains a strong irregular spiking activity (``up” state).
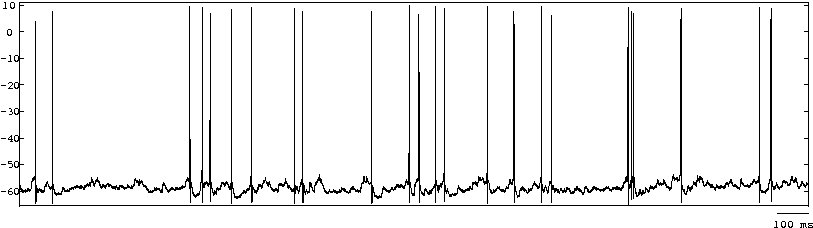
Irregularity: Oscillatory or not, it has been observed that the spiking activity of neurons in vivo is rather sparse and highly irregular. Most V1 neurons for example display Poissonian or supra-Poisson spike count variability in response to low dimensional stimuli such as bars and gratings [bibcite key=Dean1981]. This and other experimental data are in favour of the synchronous or asynchronous irregular regimes explained in the Balanced Network Section: neurons fire as Poisson sources, irregularly, with a coefficient of variation for their inter-spike intervals close to 1 [bibcite key=Nawrot2008]. The origin of this irregular activity observed in the sub-threshold voltage and/or in spiking activity is linked to synaptic activity [bibcite key=Pare1998,Destexhe1999]. To illustrate the irregularity, one can see on the left the membrane potential of a cat V1 neuron in absence of stimulation (data taken from Frégnac's lab). As we can see, the cell is spontaneously firing action potentials, and its membrane potential is fluctuating, as indirect evidence of the synaptic bombardment received by the neuron.
[/wpcol_1half_end]